兵庫県立大学 大学院生命理学研究科
細胞制御学 I 講座
兵庫県立大学 大学院生命理学研究科
細胞制御学 I 講座
研究内容
概要
体をどんなに切られても, 再生できる不死身で不思議な生き物, プラナリア. プラナリアには幹細胞が全身に存在し, 体の位置情報に従って失われた器官や組織を正しく再生することができます. 特に, ナミウズムシ (Dugesia japonica) は再生能力がとても高く, 例えば, 体を前後に切られても, 頭部からは首と腹と尾が, 尾部からは頭と首と腹が再生し, その結果, 完全な2匹のプラナリアとなります. いまから100年以上も昔に, Thomas Hunt Morgan (1933年, ショウジョウバエ遺伝学でノーベル生理学・医学賞を受賞) は, このプラナリアの再生の謎解きに挑みました. そして, モーガンは何らかの『物質の濃度勾配』が体の前後の位置情報をコードしているのではないかという仮説を提唱し, 現在においてもこの発想はあらゆる生き物の形づくりを理解するための基本概念として世界中の研究者に支持されています. そして, 我々は『モーガンの仮説』を分子レベルで解明することをめざしています.
背景
我々は, JNK蛋白質の活性化によって, 未分化なプラナリア幹細胞 (neoblast) が細胞分裂を経て, 再生芽 (blastema) を形成し, その後, ERK蛋白質の活性化によって, 様々な細胞種へと分化することを明らかにしました (図1; Tasaki et al., Dev. Growth Differ., 2011; Tasaki et al., Development, 2011). 一方で, Wnt/ß-カテニン経路が後方化シグナルとして働き, プラナリア尾部再生に必要であること(ß-カテニン遺伝子をRNA干渉法 (RNAi) によって機能阻害すると, 本来の尾部が頭部へと運命転換する) が報告されていたため, この2つのシグナル経路の組み合わせで『プラナリアの再生の仕組み』が説明できるのではないかと推測しました.
その結果, ERK蛋白質とß-カテニン蛋白質は体の前後軸に沿って相反する活性勾配を形成し, 体の異なる領域(頭, 首, 腹と尾) が再生できることを明らかにしました (図2). この仕組みにおいて, nou-darake (ノウダラケ) 遺伝子 (この遺伝子をRNAiによって機能阻害すると、脳が頭部を超えて過形成する) も関与することがわかりました. これらの結果から, プラナリアの幹細胞はERK蛋白質の活性化によって, もともと頭部の細胞に分化するように指令されますが, nou-darake遺伝子やWnt/ß-カテニン経路がERK蛋白質の活性化レベルを抑制することによって, その指令を首や腹や尾部の細胞へとそれぞれ運命転換させていると結論づけました (Umesono et al., Nature, 2013).
さらに, 我々はナミウズムシの再生原理にもとづいて, 別種のプラナリアであるコガタウズムシ (Phagocata kawakatsui: もともと尾部から頭部を再生できないプラナリア) では, Wnt/ß-カテニン経路が過剰に働くことが頭部再生不全の原因であることを実験的に証明しました (Umesono et al., Nature, 2013). ß-カテニン遺伝子をRNAiによって機能阻害されたコガタウズムシ尾部断片は, 完全に機能的な頭部を再生することができました (図3).
関連する文献
Hosoda, K., Motoishi, M., Kunimoto, T., Nishimura, O., Hwang, B., Kobayashi, S., Yazawa, S., Mochii, M., Agata, K. and Umesono, Y. (2018). Role of MEKK1 in the anterior-posterior patterning during planarian regeneration. Dev. Growth Differ., 60: 341-353.
---------------------------------------------------------------------------------------------------------------------------------------
Umesono, Y., Tasaki, J., Nishimura, Y., Hrouda, M., Kawaguchi, E., Yazawa, S., Nishimura, O., Hosoda, K., Inoue, T. and Agata, K. (2013). The molecular logic for planarian regeneration along the anterior–posterior axis. Nature, 500: 73-76.
---------------------------------------------------------------------------------------------------------------------------------------
Tasaki, J., Shibata, N., Nishimura, O., Itomi, K., Tabata, Y., Son, F., Suzuki, N., Araki, R., Abe, M., Agata, K. and Umesono, Y. (2011). ERK signaling controls blastema cell differentiation during planarian regeneration. Development, 138: 2417-2427.
---------------------------------------------------------------------------------------------------------------------------------------
Tasaki, J., Shibata, N., Sakurai, T., Agata, K. and Umesono, Y. (2011). Role of c-Jun N-terminal kinase activation in blastema formation during planarian regeneration. Dev. Growth Differ., 53: 389-400.
---------------------------------------------------------------------------------------------------------------------------------------
Yazawa, S., Umesono, Y., Hayashi, T., Tarui, H. and Agata, K. (2009). Planarian Hedgehog/Patched establishes anterior-posterior polarity by regulating Wnt signaling. Proc. Natl. Acad. Sci. USA, 106: 22329-22334.
---------------------------------------------------------------------------------------------------------------------------------------
Cebrià, F., Kobayashi, C., Umesono, Y., Nakazawa, M., Mineta, K., Ikeo, K., Gojobori, T., Ito, M., Taira, M., Sánchez Alvarado, A. and Agata, K. (2002). FGFR-related gene nou-darake restricts brain tissues to the head region of planarians. Nature, 419: 620-624.
---------------------------------------------------------------------------------------------------------------------------------------
研究課題(現在進行中_1)
ERK経路活性化に関わる上流因子群の同定.
研究課題(現在進行中_2)
nou-darake遺伝子やWnt/ß-カテニン経路がERK経路活性化を抑制する制御機構の解明.
研究課題(現在進行中_3)
プラナリア再生過程における前後軸に沿ったからだのプロポーションを制御している分子機構の解明.

図2:プラナリアの再生原理.
ERK活性化レベルは頭部で最も高く, 首部ではnou-darake遺伝子によって低くなっている. 腹部および尾部では, ERK活性化レベルはß-カテニン活性によってさらに低くなり, 頭尾軸に沿った勾配を形成していると考えられる. nou-darake遺伝子およびß-カテニン遺伝子をノックダウンすると、ERK活性化レベルの上昇により勾配はなくなり, すべての領域が頭部に運命転換する.
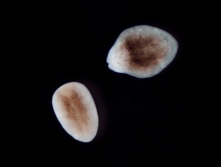
図3:コガタウズムシ尾部断片からの頭部再生誘導.
左図は正常な尾部断片. 頭部を再生することができない.
右図はß-カテニン遺伝子をノックダウンした尾部断片.
白い部分が再生した頭部. 1対の眼が確認できる.
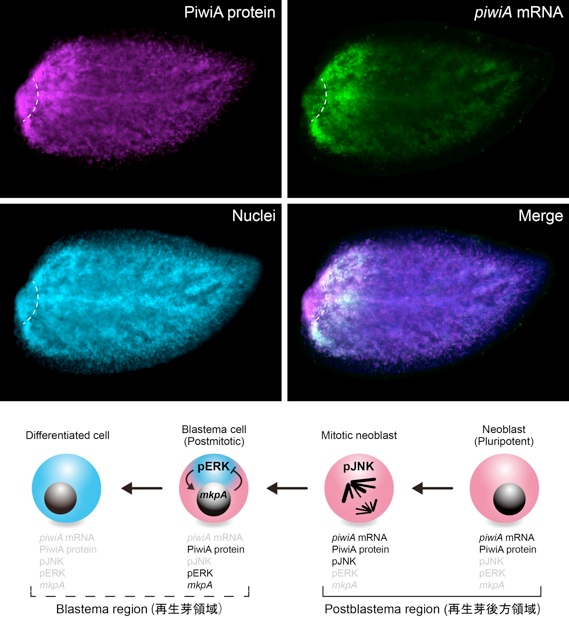
図1:プラナリア幹細胞 (neoblast) の分化制御機構.

